
Открытие.
Структуру, известную как комплекс Гольджи, впервые обнаружил в клетках
животных в 1898 г. Камилло Гольджи, итальянский врач и цитолог. Детальное исследование данной структуры сделано позже с помощью электронного микроскопа.
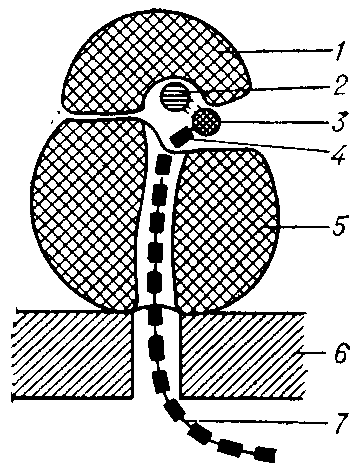
Аппарат Гольджи содержится в цитоплазме почти всех эукариотических клеток, особенно в секреторных клетках животных. У дрожжей комплекс Гольджи выражен несколько хуже, обычно в виде особого отдела эндоплазматического ретикулума. Комплекс Гольджи представляет собой стопку уплощенных мембранных мешочков, так называемых цистерн, и связанную с ними систему пузырьков, называемых пузырьками Гольджи. На одном конце стопки
мешочков постоянно образуются новые цистерны путем слияния пузырьков, отпочковывающихся, от гладкого эндоплазматического ретикулума. На другом конце стопки, на внутренней стороне завершается созревание цистерн и они вновь распадаются на пузырьки. Таким образом, цистерны в стопке постепенно перемещаются от наружной стороны к внутренней.
Функцией аппарата Гольджи является транспорт и химиче
ская модификация поступающих в него веществ. Исходным субстратом для ферментов являются белки, поступающие в аппарат Гольджи из эндоплазматического ретикулума. После модификации и концентрирования, ферменты в пузырьках Гольджи переносятся к «месту назначения», например к месту образования новой почки. Наиболее активно этот перенос осуществляется с участием цитоплазматических микротрубочек.
Аппарат Гольджи является компонентом всех эукариотических клеток (практически единственное исключение - эритроциты млекопитающих). Он п
редставляет собой важнейшую мембранную органеллу, управляющую процессами внутриклеточного транспорта. Основными функциями аппарата Гольджи являются модификация, накопление, сортировка и направление различных веществ в соответствующие внутриклеточные компартменты, а также за пределы клетки. Он состоит из набора окруженных мембраной уплощенных цистерн , напоминающих стопку тарелок. Каждая стопка Гольджи (у растений называемая диктиосомой ) обычно содержит от четырех до шести цистерн, имеющих, как правило, диаметр около 1мкм ( рис. 8-36 ). Число стопок Гольджи в клетке в значительной степени зависит от ее типа: некоторые клетки содержат одну большую стопку, тогда как в других имеются сотни очень маленьких стопок.
Со стопками Гольджи всегда ассоциирована масса мелких (диаметром приблизительно 60 нм) ограниченных мембраной пузырьков. Полагают,
что эти пузырьки (пузырьки Гольджи ) переносят белки и липиды в аппарат Гольдки, транспортируют их из него и между остальными цистернами. Многие пузырьки являются окаймленными и покрыты клатрином или другим специфическим белком. Часто можно видеть, как такие окаймленные пузырьки отшнуровываются от цистерн Гольджи.
Аппарат Гольджи имеет две разные стороны: формирующуюся, или цис-сторону и зрелую, или транс-сторону Цис-сторона тесно связана с переходными элементами ЭР; транс-сторона расширяется, образуя трубчатый ретикулум, называемый транс-сетью Гольджи . Белки и липиды в составе небольших пузырьков попадают в стопку Гольджи с цис-стороны, а покидают ее, направляясь в различные компартменты, вместе с пузырьками, образующимися на транс-стороне. Переходя из одной стопки Гольдж
и в другую, эти молекулы претерпевают последовательные серии модификаций.
Структура.
Описание структуры аппарата Гольджи тесно связано с описанием его основных биохимических функций, поскольку подразделение этогоклеточного

компартмента на отделы производится преимущественно на основе локализации ферментов, расположенных в том или ином отделе.
Чаще всего в аппарате Гольджи выделяют четыре основных отдела: цис- Гольджи , медиал-Гольджи , транс-Гольджи и транс-Гольджи сеть ( TGN )
Кроме того к аппарату Гольджи иногда относят так называемыйпромежуточный компартмент, представляющий собой скопление мембранных пузырьков между эндоплазматическим ретикулумом и цис-Гольджи. Аппарат Гольджи является очень полиморфной органеллой; в клетках разных типов и даже на разных стадиях развития одной и той же клетки он может выглядеть по-разному. Основные его характеристики таковы:
- наличие стопки из нескольких (обычно 3-8) уплощенных цистерн, более или менее плотно прилегающих друг к другу. Такая стопка всегда бывает окружена некоторым (иногда очень значительным) количеством мембранных пузырьков. В животных клетках чаще можно встретить одну стопку, в то время как в растительных клетках их обычно бывает несколько; каждую из них в таком случае называют диктиосомой. Отдельные диктиосомы могут быть связаны между собой системой вакуолей, образуя трехмерную сеть;
- композиционная гетерогенность, выражающаяся в том, что постоянные (resident) ферменты неоднородно распределены по органелле;
- полярность, то есть наличие цис-стороны, обращенной к эндоплазматическому ретикулуму и ядру, и транс-стороны,обращенной к поверхности клетки (это особенно характерно для секретирующих клеток);
- ассоциация с микротрубочками и областью центриоли. Разрушение микротрубочек деполимеризующими агентами приводит к фрагментации аппарата Гольджи, однако его функции при этом существенно не затрагиваются. Аналогичная фрагментация наблюдается и в естественных условиях, во время митоза . После восстановления системы микротрубочек разбросанные по клетке элементы аппарата Гольджи собираются (по микротру-бочкам) в область центриоли,и реконструируется нормальный комплекс Гольджи.
Аппарат Гольджи (комплекс Гольджи) — мембранная структура эукариотической клетки, в основном предназначенная для выведения веществ, синтезированных в эндоплазматическом ретикулуме. Комплекс Гольджи был назван так в честь итальянского ученого Камилло Гольджи, впервые обнаружившего его в 1898 году.
Комплекс Гольдки представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям и связанную с ними систему пузырьков Гольдки. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько соединенных трубками стопок.
В цистернах Аппарата Гольджи созревают белки предназначенные для секреции, трансмембранные белки плазматической мембраны, белки лизосом и т.д. Созревающие белки последовательно перемещаются по цистернам органеллы, в которых происходит их окончательное сворачивание, а также модификации — гликозилирование и фосфорилирование.
Аппарат Гольджи ассиметричен — цистерны располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от гранулярного эндоплазматического ретикулума (ЭР), на мембранах которого и происходит синтез белков рибосомами.
Разные цистерны Аппарата Гольджи содержат разные резидентные каталитические ферменты и, следовательно, с созревающими белками в них последовательно происходят разные процессы. Понятно, что такой ступенчатый процесс должен как-то контролироваться. Действительно, созревающие белки «маркируются» специальными полисахаридными остатками (преимущественно маннозными), по-видимому, играющими роль своебразного «знака качества».
Не до конца понятно, каким образом созревающие белки перемещаются по цистернам Аппарата Гольджи, в то время как резидентные белки остаются в большей или меньшей степени ассоциированы с одной цистерной. Существуют две взаимонеисключающие гипотезы, объясняющие этот механизм. Согласно первой (1), транспорт белков осуществляется при помощи таких же механизмов везикулярного транспорта, как и путь транспорта из ЭР, причем резидентные белки не включаются в отпочковывающуюся везикулу. Согласно второй (2), происходит непрерывное передвижение (созревание) самих цистерн, их сборка из пузырьков с одного конца и разборка с другого конца органеллы, а резидентные белки перемещаются ретроградно (в обратном направлении) при помощи везикулярного транспорта.
В конце концов от противоположного конца органеллы (транс-Гольджи) отпочковываются пузырьки, содержащие полностью зрелые белки.
В комплексе Гольджи происходит
1. О-гликозилирование, к белкам присоединяются сложные сахара через атом кислорода.
2. Фосфорилирование (присоединение к белкам остатка ортофосфорной кислоты).
3. Образование лизосом.
4. Образование клеточной стенки (у растений).
5. Участие в везикулярном транспорте (формирование трехбелкового потока):
6. созревание и транспорт белков плазматической мембраны;
7. созревание и транспорт секретов;
8. созревание и транспорт ферментов лизосом.
Аппарат Гольджи (комплекс Гольджи) – это специализированная часть эндоплазматического ретикулума, состоящая из собранных в стопки плоских мембранных мешочков. Он участвует в секреции клеткой белков (в нем происходит упаковка секретируемых белков в гранулы) и поэтому особенно развит в клетках, выполняющих секреторную функцию. К важным функциям аппарата Гольдки относится также присоединение углеводных групп к белкам и использование этих белков для построения клеточной мембраны и мембраны лизосом. У некоторых водорослей в аппарате Гольджи осуществляется синтез волокон целлюлозы.
Функции.
Функцией аппарата Гольджи является транспорт и химическая модификация поступающих в него веществ. Исходным субстратом для ферментов являются белки, поступающие в аппарат Гольджи из эндоплазматического ретикулума. После модификации и концентрирования, ферменты в пузырьках Гольджи переносятся к «месту назначения», например к месту образования новой почки. Наиболее активно этот перенос осуществляется с участием цитоплазматических микротрубочек.
Функции аппарата Гольджи очень многообразны. К ним можно отнести:
1) сортировку, накопление и выведение секреторных продуктов;
2) завершение посттрансляционной модификации белков ( гликозилирование , сульфатирование и т.д.);
3) накопление молекул липидов и образование липопротеидов ;
4) образование лизосом ;
5) синтез полисахаридов для образования гликопротеидов, восков, камеди, слизей, веществ матрикса клеточных стенок растений
(гемицеллюлоза, пектины) и т.п.
6) формирование клеточной пластинки после деления ядра в растительных клетках;
7) участие в формировании акросомы ;
8) формирование сократимых вакуолей простейших.
Этот список, без сомнения, неполон, и дальнейшие исследования не только позволят лучше понять уже известные функции аппарата Гольджи, но и приведут к открытию новых. Пока самыми изученными с биохимической точки зрения остаются функции, связаные с транспортом и модификацией новосинтезированных белков.
 Клетка бактерии
Клетка бактерии
 Строение прокариотической клетки
Строение прокариотической клетки







 Вы видите, что мембрана образована двумя слоями липидов, а белковые молекулы пронизывают толщу мембраны.Основная функция плазматической мембраны транспортная. Она обеспечивает поступление питательных веществ в клетку и выведение из нее продуктов обмена.Важное свойство мембраны – избирательная проницаемость, или полупроницаемость, позволяет клетке взаимодействовать с окружающей средой: в нее поступают и выводятся из нее лишь определенные вещества
Вы видите, что мембрана образована двумя слоями липидов, а белковые молекулы пронизывают толщу мембраны.Основная функция плазматической мембраны транспортная. Она обеспечивает поступление питательных веществ в клетку и выведение из нее продуктов обмена.Важное свойство мембраны – избирательная проницаемость, или полупроницаемость, позволяет клетке взаимодействовать с окружающей средой: в нее поступают и выводятся из нее лишь определенные вещества Липиды в мембране образуют двойной слой, а белки пронизывают всю ее толщу,погружены на разную глубину в липидный слой или располагаются на внешней ивнутренней поверхности мембраны.К некоторым белкам, находящимся на наружной поверхности, прикреплеплены углеводы. Белки и углеводы на поверхности мембран у разных клеток неодинаковы и являются своеобразным указателем типа клеток.Например,с помощью этих указателей сперматазоиды узнают яйцеклетку.Благодаря мембранным "антеннам" клетки,принадлежащие к одному типу,удерживаются вместе,образуя ткани.Строение мембран всех других органоидов сходно с плазматической мембраной. Различаются они составом, соотношением липидов и белков, их расположением в структуре мембран.
Липиды в мембране образуют двойной слой, а белки пронизывают всю ее толщу,погружены на разную глубину в липидный слой или располагаются на внешней ивнутренней поверхности мембраны.К некоторым белкам, находящимся на наружной поверхности, прикреплеплены углеводы. Белки и углеводы на поверхности мембран у разных клеток неодинаковы и являются своеобразным указателем типа клеток.Например,с помощью этих указателей сперматазоиды узнают яйцеклетку.Благодаря мембранным "антеннам" клетки,принадлежащие к одному типу,удерживаются вместе,образуя ткани.Строение мембран всех других органоидов сходно с плазматической мембраной. Различаются они составом, соотношением липидов и белков, их расположением в структуре мембран.


{kind=link}